Variability and directionality of inferior olive neuron dendrites revealed by detailed 3D characterization of an extensive morphological library
- PMID: 30929054
- PMCID: PMC6509097
- DOI: 10.1007/s00429-019-01859-z
Variability and directionality of inferior olive neuron dendrites revealed by detailed 3D characterization of an extensive morphological library
Abstract
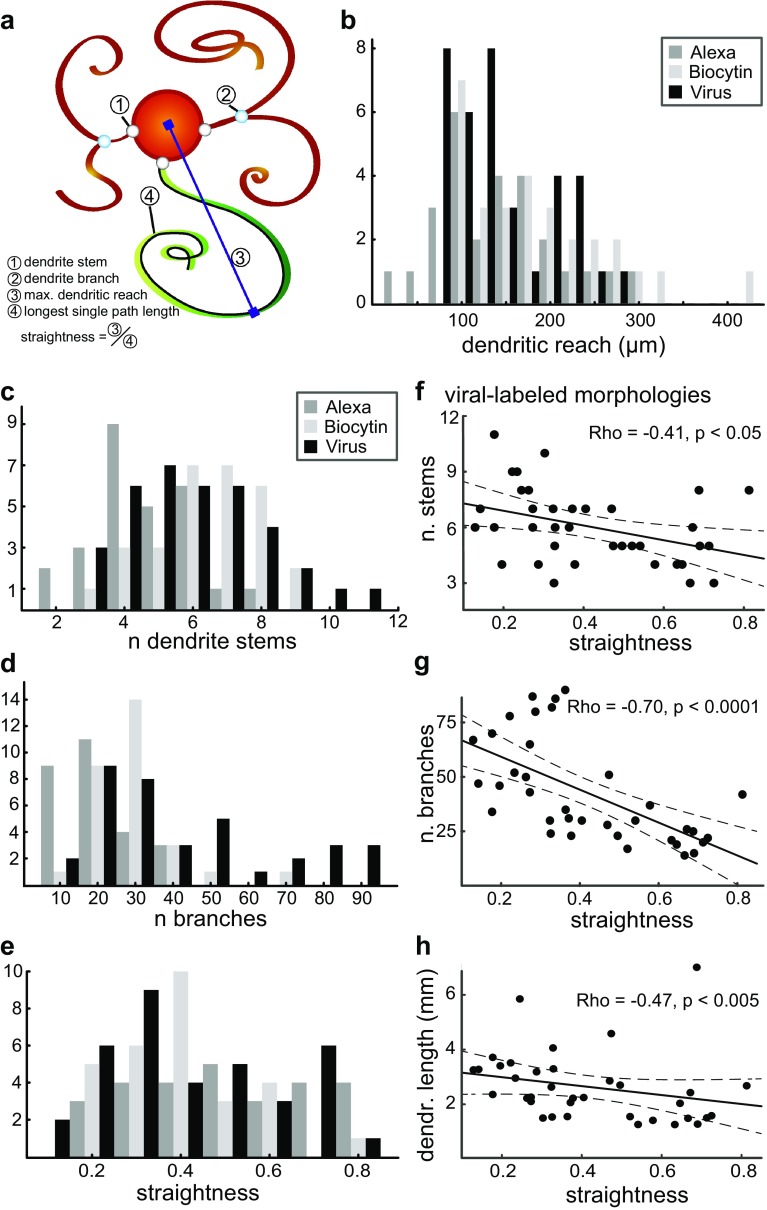
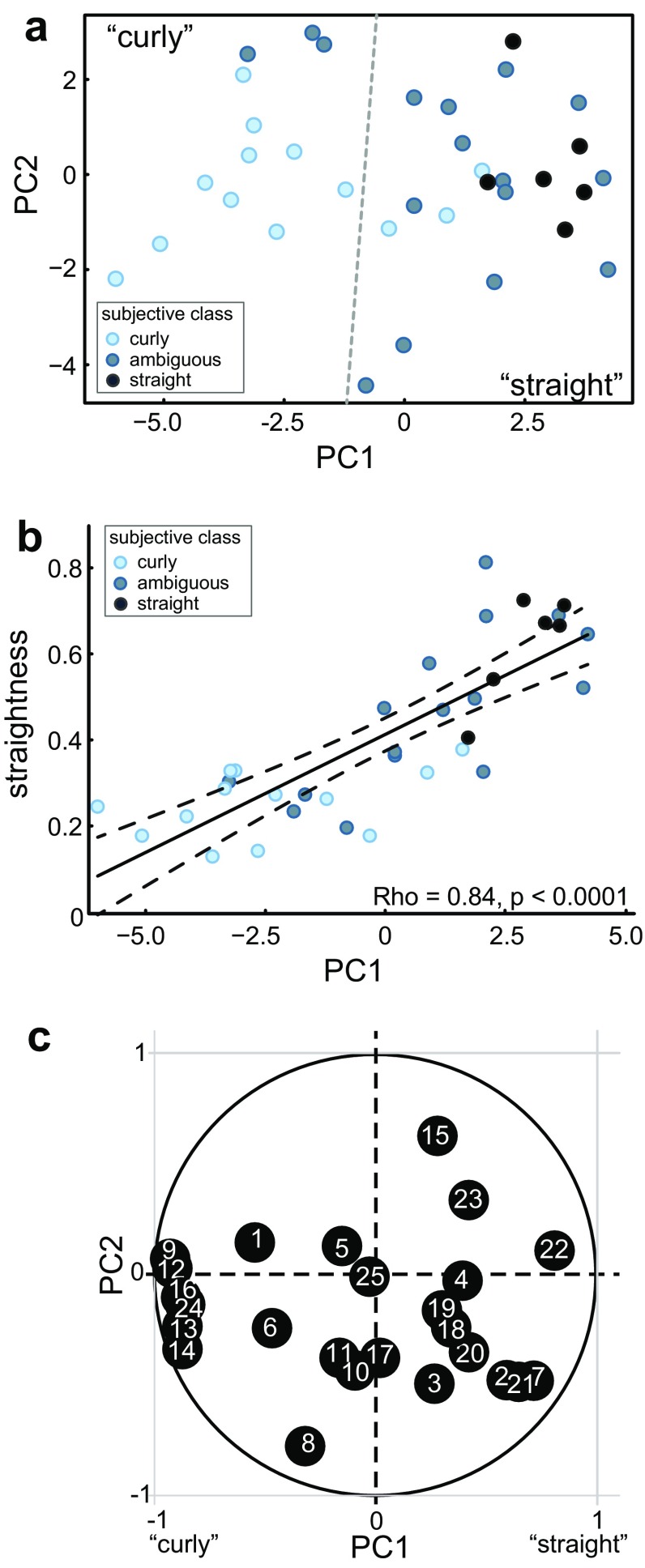
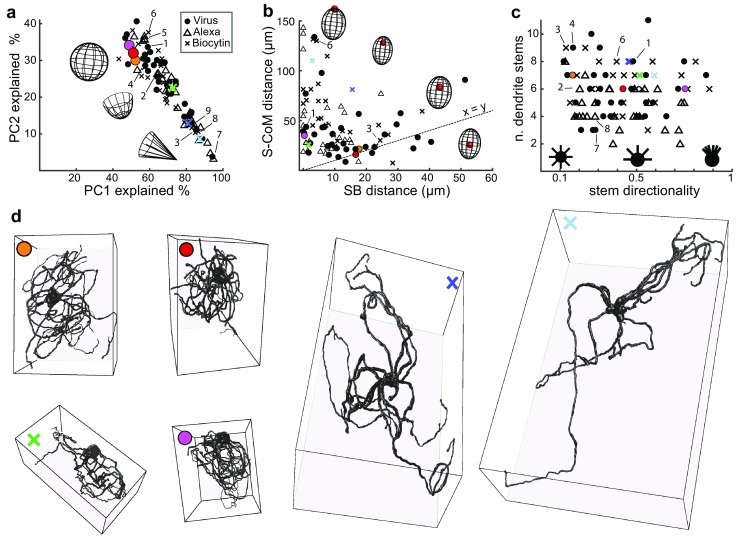
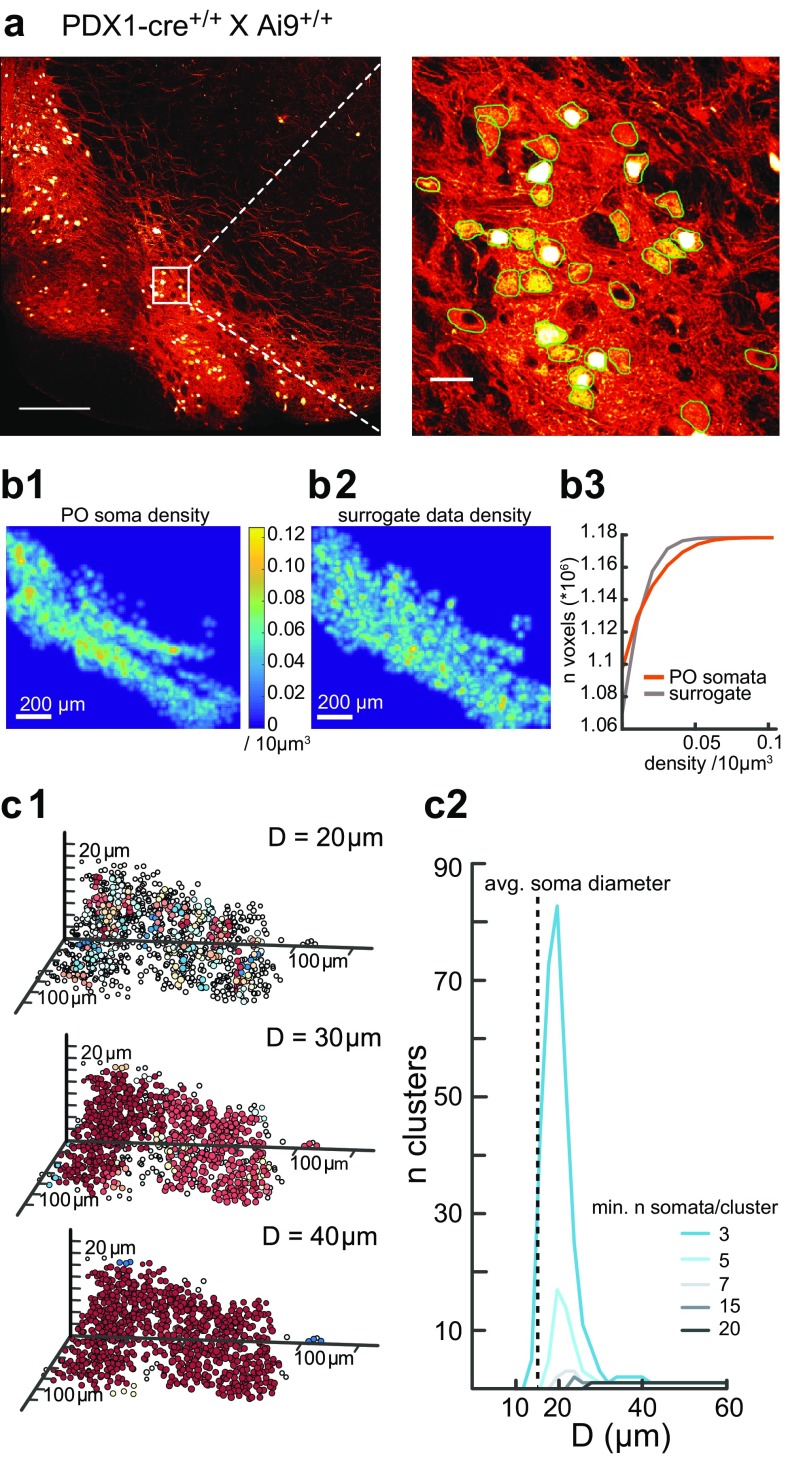
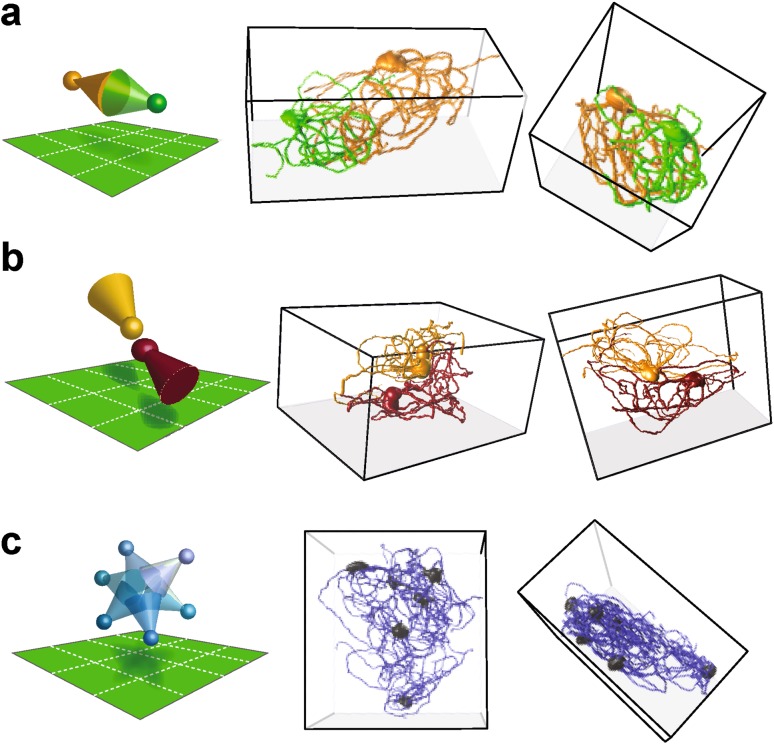
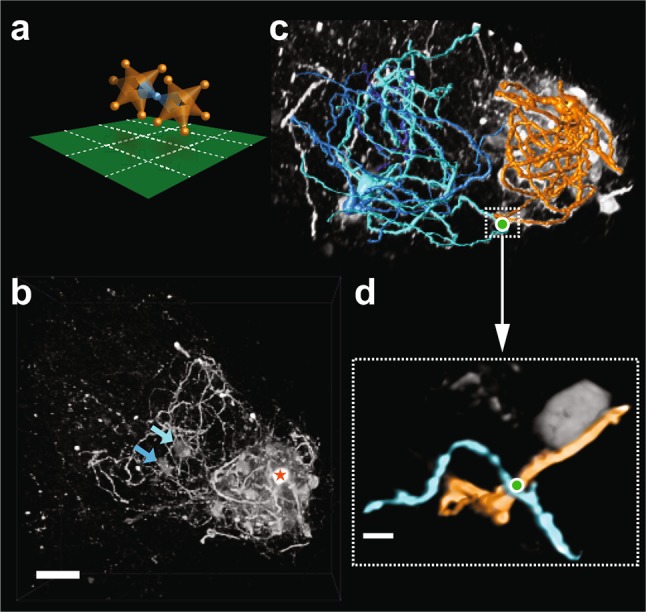
The inferior olive (IO) is an evolutionarily conserved brain stem structure and its output activity plays a major role in the cerebellar computation necessary for controlling the temporal accuracy of motor behavior. The precise timing and synchronization of IO network activity has been attributed to the dendro-dendritic gap junctions mediating electrical coupling within the IO nucleus. Thus, the dendritic morphology and spatial arrangement of IO neurons governs how synchronized activity emerges in this nucleus. To date, IO neuron structural properties have been characterized in few studies and with small numbers of neurons; these investigations have described IO neurons as belonging to two morphologically distinct types, "curly" and "straight". In this work we collect a large number of individual IO neuron morphologies visualized using different labeling techniques and present a thorough examination of their morphological properties and spatial arrangement within the olivary neuropil. Our results show that the extensive heterogeneity in IO neuron dendritic morphologies occupies a continuous range between the classically described "curly" and "straight" types, and that this continuum is well represented by a relatively simple measure of "straightness". Furthermore, we find that IO neuron dendritic trees are often directionally oriented. Combined with an examination of cell body density distributions and dendritic orientation of adjacent IO neurons, our results suggest that the IO network may be organized into groups of densely coupled neurons interspersed with areas of weaker coupling.
Keywords: Brainstem; Dendritic morphometry; Network structure; Neuron reconstructions; Olivo-cerebellar system; Sparse viral labeling.
Conflict of interest statement
Conflict of interest
The authors declare that they have no conflict of interest.
Research involving human participants and/or animals
No human participants. All animal experimental procedures were approved by the Hebrew University’s Animal Care and Use Committee, and the animal experiment committee of the Royal Netherlands Academy of Arts and Sciences (DEC-KNAW) which follows the European guidelines for the care and use of laboratory animals (Council Directive 86/6009/EEC).
Informed consent
Not applicable.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous