Flexible timing by temporal scaling of cortical responses
- PMID: 29203897
- PMCID: PMC5742028
- DOI: 10.1038/s41593-017-0028-6
Flexible timing by temporal scaling of cortical responses
Abstract
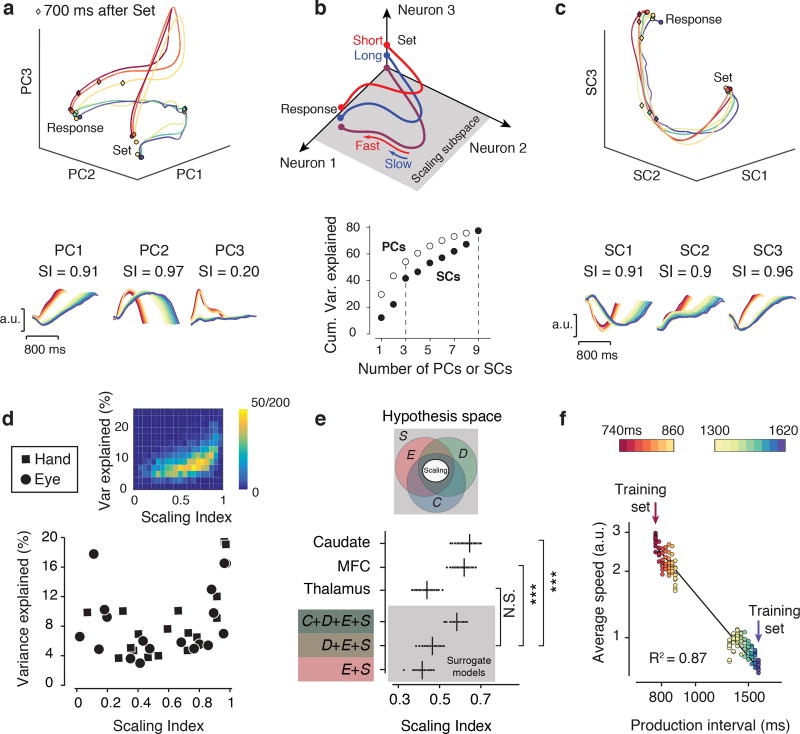
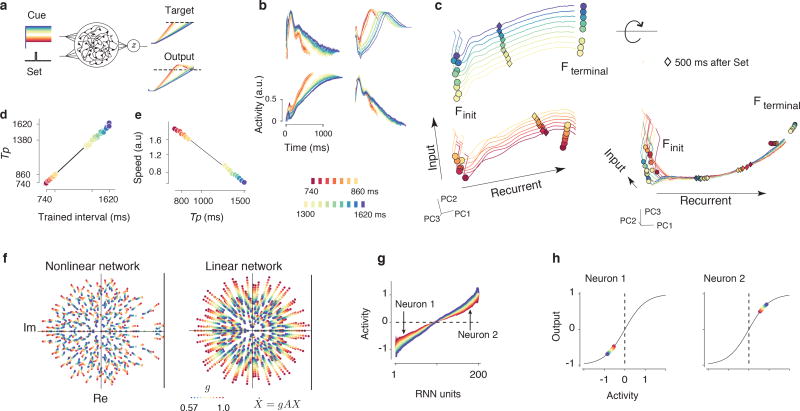
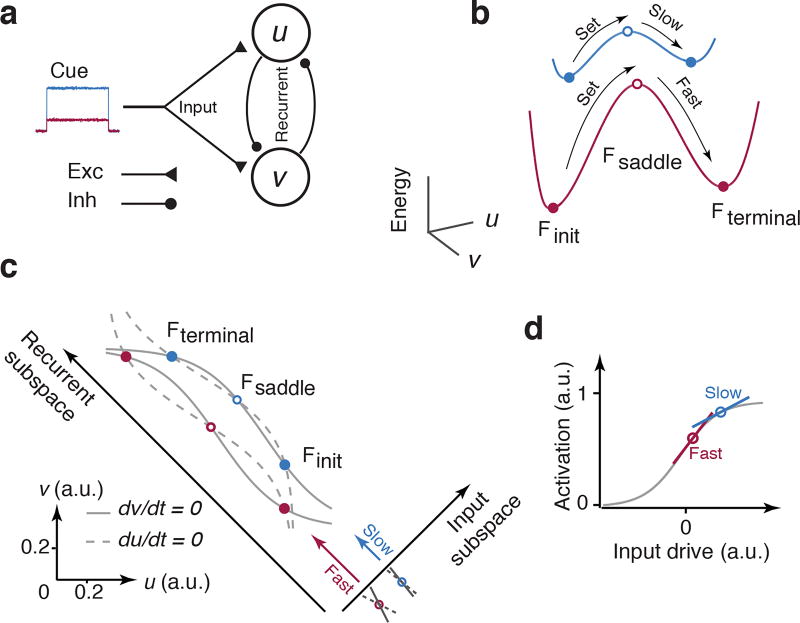
Musicians can perform at different tempos, speakers can control the cadence of their speech, and children can flexibly vary their temporal expectations of events. To understand the neural basis of such flexibility, we recorded from the medial frontal cortex of nonhuman primates trained to produce different time intervals with different effectors. Neural responses were heterogeneous, nonlinear, and complex, and they exhibited a remarkable form of temporal invariance: firing rate profiles were temporally scaled to match the produced intervals. Recording from downstream neurons in the caudate and from thalamic neurons projecting to the medial frontal cortex indicated that this phenomenon originates within cortical networks. Recurrent neural network models trained to perform the task revealed that temporal scaling emerges from nonlinearities in the network and that the degree of scaling is controlled by the strength of external input. These findings demonstrate a simple and general mechanism for conferring temporal flexibility upon sensorimotor and cognitive functions.
Conflict of interest statement
The authors declare no conflicting interests.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Stuphorn V, Schall JD. Executive control of countermanding saccades by the supplementary eye field. Nat. Neurosci. 2006;9:925–931. - PubMed
-
- Kunimatsu J, Tanaka M. Alteration of the timing of self-initiated but not reactive saccades by electrical stimulation in the supplementary eye field. Eur. J. Neurosci. 2012;36:3258–3268. - PubMed
-
- Lewis PA, Wing AM, Pope PA, Praamstra P, Miall RC. Brain activity correlates differentially with increasing temporal complexity of rhythms during initialisation, synchronisation, and continuation phases of paced finger tapping. Neuropsychologia. 2004;42:1301–1312. - PubMed
-
- Shima K, Tanji J. Neuronal activity in the supplementary and presupplementary motor areas for temporal organization of multiple movements. J. Neurophysiol. 2000;84:2148–2160. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources