Neural mechanisms of speed-accuracy tradeoff
- PMID: 23141072
- PMCID: PMC3576837
- DOI: 10.1016/j.neuron.2012年08月03日0
Neural mechanisms of speed-accuracy tradeoff
Abstract
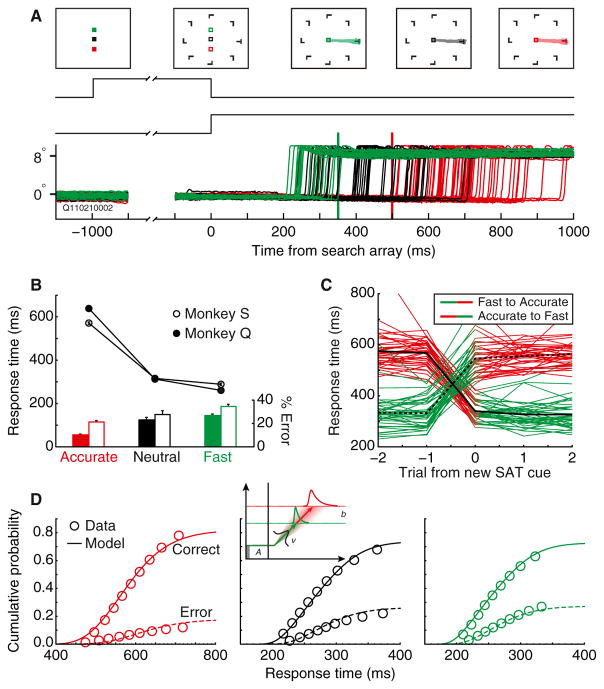
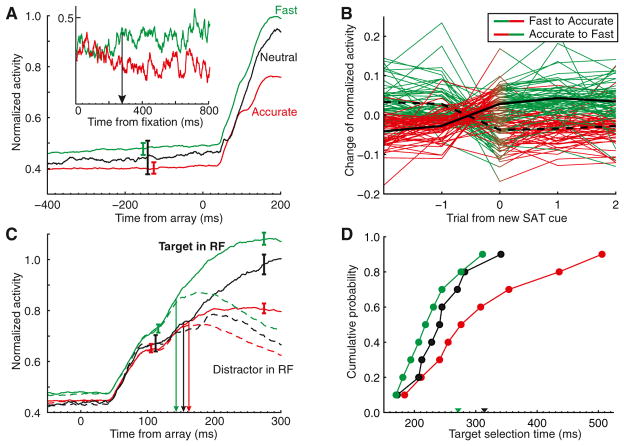
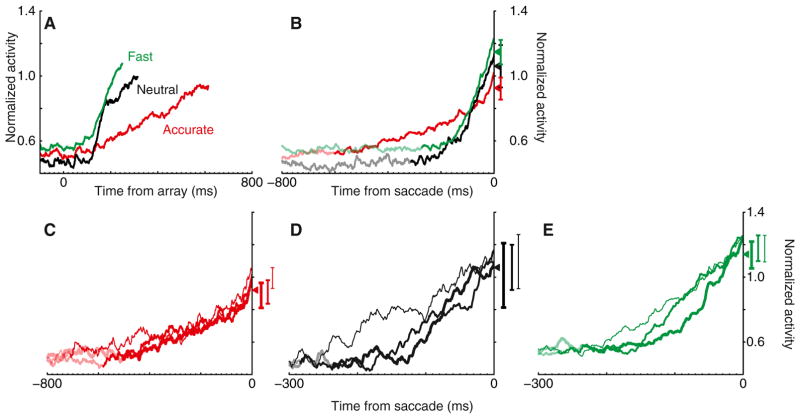
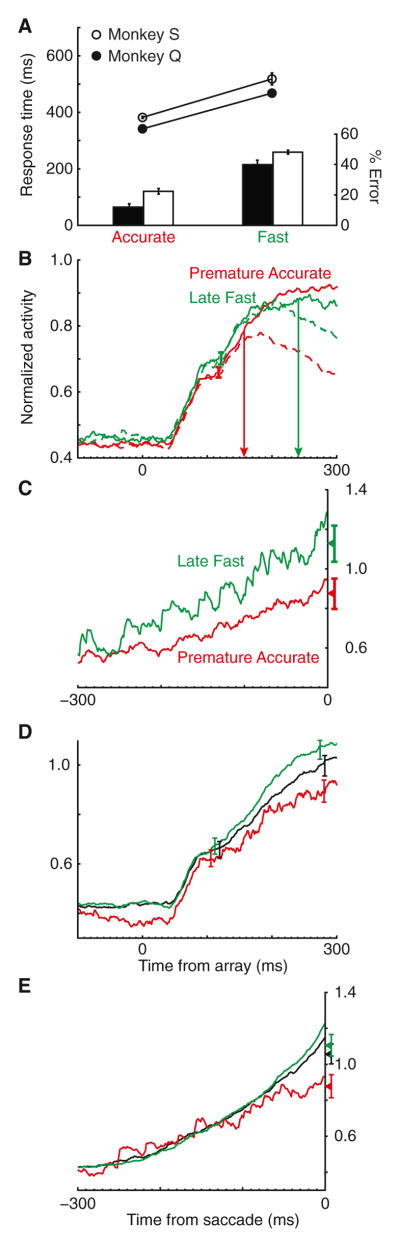
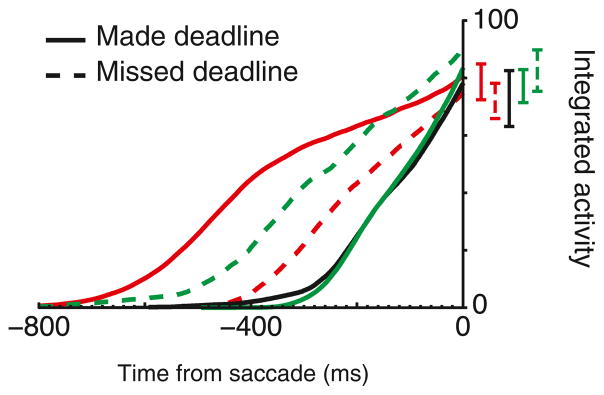
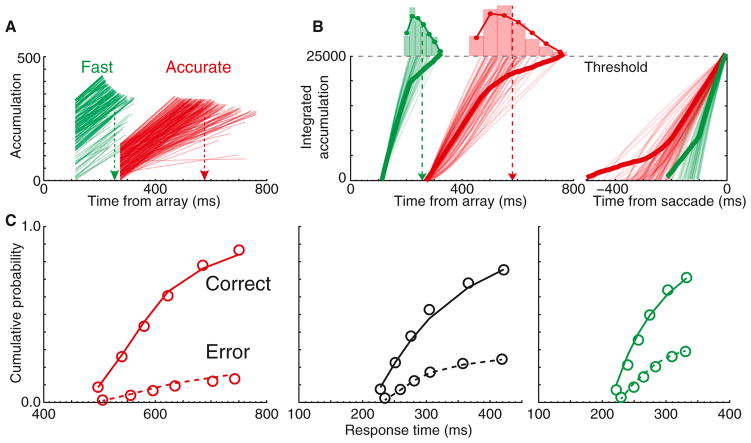
Intelligent agents balance speed of responding with accuracy of deciding. Stochastic accumulator models commonly explain this speed-accuracy tradeoff by strategic adjustment of response threshold. Several laboratories identify specific neurons in prefrontal and parietal cortex with this accumulation process, yet no neurophysiological correlates of speed-accuracy tradeoff have been described. We trained macaque monkeys to trade speed for accuracy on cue during visual search and recorded the activity of neurons in the frontal eye field. Unpredicted by any model, we discovered that speed-accuracy tradeoff is accomplished through several distinct adjustments. Visually responsive neurons modulated baseline firing rate, sensory gain, and the duration of perceptual processing. Movement neurons triggered responses with activity modulated in a direction opposite of model predictions. Thus, current stochastic accumulator models provide an incomplete description of the neural processes accomplishing speed-accuracy tradeoffs. The diversity of neural mechanisms was reconciled with the accumulator framework through an integrated accumulator model constrained by requirements of the motor system.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Bogacz R, Brown E, Moehlis J, Holmes P, Cohen JD. The physics of optimal decision making: a formal analysis of models of performance in two-alternative forced-choice tasks. Psychol Rev. 2006;113:700–765. - PubMed
-
- Bogacz R, Wagenmakers EJ, Forstmann BU, Nieuwenhuis S. The neural basis of the speed-accuracy tradeoff. Trends Neurosci. 2010;33:10–16. - PubMed
-
- Boucher L, Palmeri TJ, Logan GD, Schall JD. Inhibitory control in mind and brain: an interactive race model of countermanding saccades. Psychol Rev. 2007;114:376–397. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources