Differential olivo-cerebellar cortical control of rebound activity in the cerebellar nuclei
- PMID: 20395550
- PMCID: PMC2889566
- DOI: 10.1073/pnas.0907118107
Differential olivo-cerebellar cortical control of rebound activity in the cerebellar nuclei
Abstract
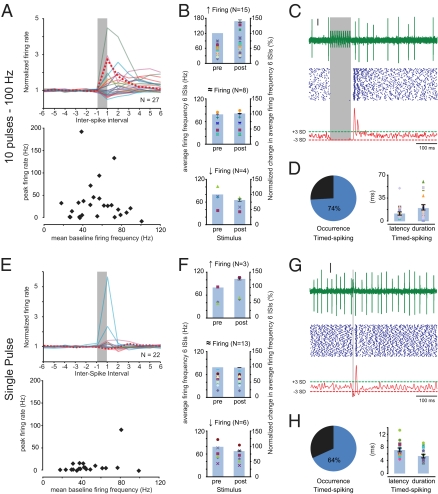
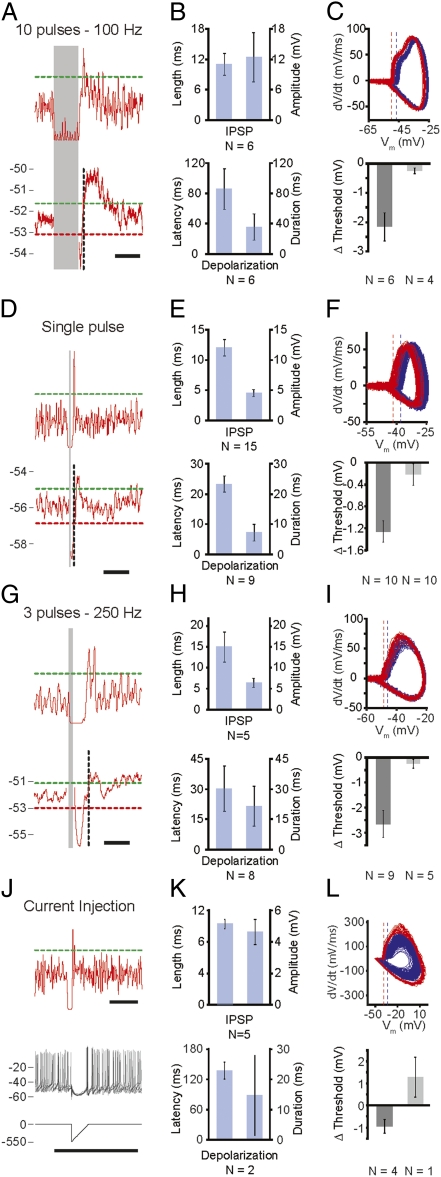
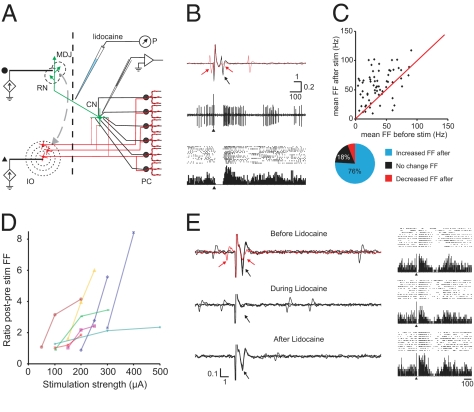
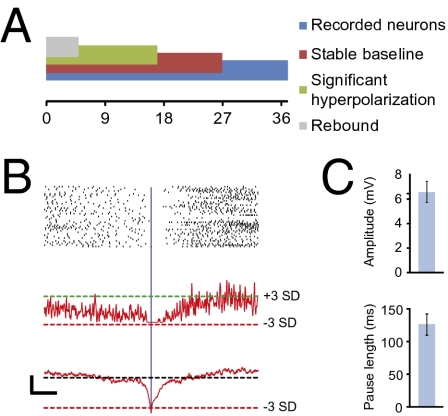
The output of the cerebellar cortex is controlled by two main inputs, (i.e., the climbing fiber and mossy fiber-parallel fiber pathway) and activations of these inputs elicit characteristic effects in its Purkinje cells: that is, the so-called complex spikes and simple spikes. Target neurons of the Purkinje cells in the cerebellar nuclei show rebound firing, which has been implicated in the processing and storage of motor coordination signals. Yet, it is not known to what extent these rebound phenomena depend on different modes of Purkinje cell activation. Using extracellular as well as patch-clamp recordings, we show here in both anesthetized and awake rodents that simple and complex spike-like train stimuli to the cerebellar cortex, as well as direct activation of the inferior olive, all result in rebound increases of the firing frequencies of cerebellar nuclei neurons for up to 250 ms, whereas single-pulse stimuli to the cerebellar cortex predominantly elicit well-timed spiking activity without changing the firing frequency of cerebellar nuclei neurons. We conclude that the rebound phenomenon offers a rich and powerful mechanism for cerebellar nuclei neurons, which should allow them to differentially process the climbing fiber and mossy fiber inputs in a physiologically operating cerebellum.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- De Zeeuw CI, Berrebi AS. Postsynaptic targets of Purkinje cell terminals in the cerebellar and vestibular nuclei of the rat. Eur J Neurosci. 1995;7:2322–2333. - PubMed
-
- Teune TM, van der Burg J, van der Moer J, Voogd J, Ruigrok TJ. Topography of cerebellar nuclear projections to the brain stem in the rat. Prog Brain Res. 2000;124:141–172. - PubMed
-
- Uusisaari M, Knöpfel T. GABAergic synaptic communication in the GABAergic and non-GABAergic cells in the deep cerebellar nuclei. Neuroscience. 2008;156:537–549. - PubMed
-
- Chan-Palay V. Cerebellar Dentate Nucleus: Organization, Cytology and Transmitters. Berlin: Springer-Verlag; 1977.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources