Inward-facing conformation of glutamate transporters as revealed by their inverted-topology structural repeats
- PMID: 19926849
- PMCID: PMC2791632
- DOI: 10.1073/pnas.0908570106
Inward-facing conformation of glutamate transporters as revealed by their inverted-topology structural repeats
Abstract
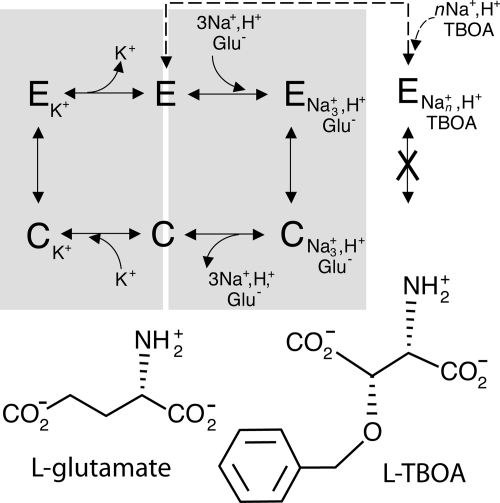
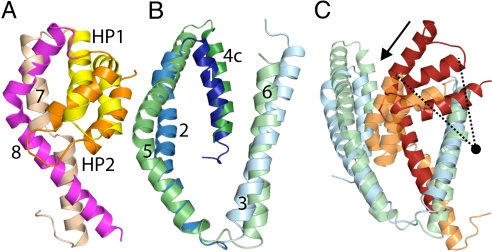
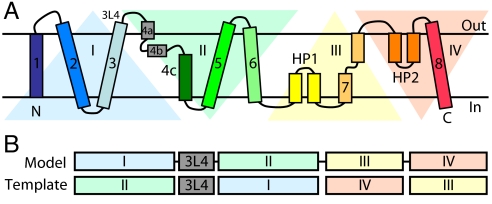
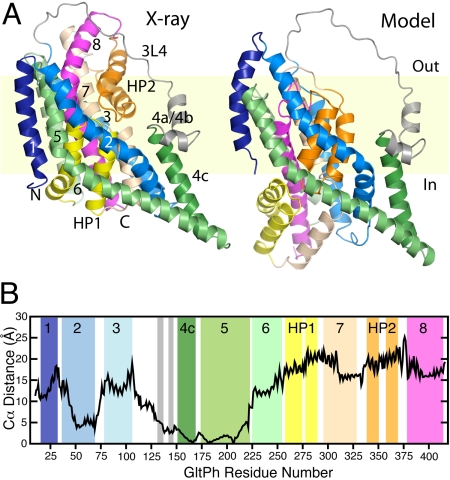
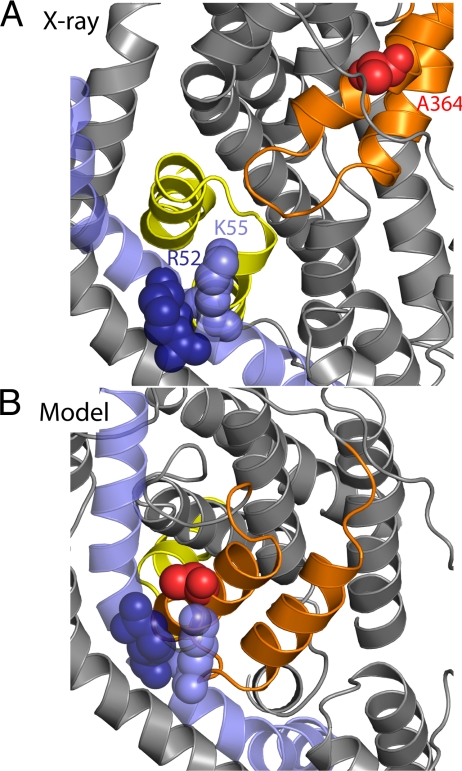
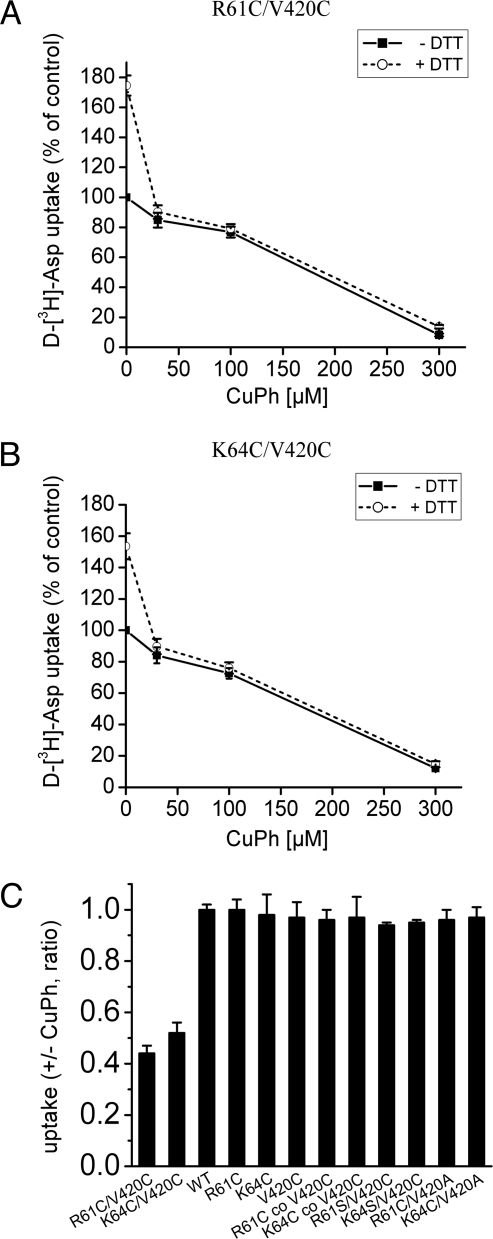
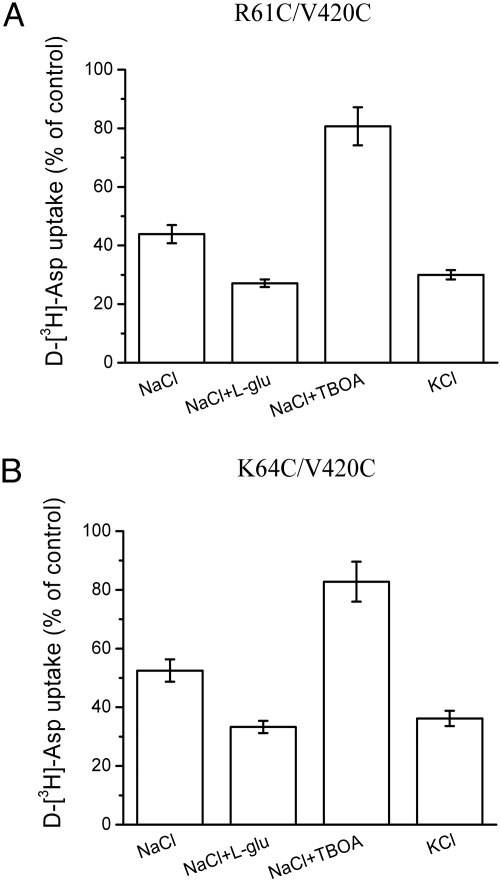
Glutamate transporters regulate synaptic concentrations of this neurotransmitter by coupling its flux to that of sodium and other cations. Available crystal structures of an archeal homologue of these transporters, GltPh, resemble an extracellular-facing state, in which the bound substrate is occluded only by a small helical hairpin segment called HP2. However, a pathway to the cytoplasmic side of the membrane is not clearly apparent. We previously modeled an alternate state of a transporter from the neurotransmitter:sodium symporter family, which has an entirely different fold, solely on the presence of inverted-topology structural repeats. In GltPh, we identified two distinct sets of inverted-topology repeats and used these repeats to model an inward-facing conformation of the protein. To test this model, we introduced pairs of cysteines into the neuronal glutamate transporter EAAC1, at positions that are >27 A apart in the crystal structures of GltPh, but approximately = 10 A apart in the inward-facing model. Transport by these mutants was activated by pretreatment with the reducing agent dithithreitol. Subsequent treatment with the oxidizing agent copper(II)(1,10-phenantroline)(3) abolished this activation. The inhibition of transport was potentiated under conditions thought to promote the inward-facing conformation of the transporter. By contrast, the inhibition was reduced in the presence of the nontransportable substrate analogue D,L-threo-beta-benzyloxyaspartate, which favors the outward-facing conformation. Other conformation-sensitive accessibility measurements are also accommodated by our inward-facing model. These results suggest that the inclusion of inverted-topology repeats in transporters may provide a general solution to the requirement for two symmetry-related states in a single protein.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Zerangue N, Kavanaugh MP. Flux coupling in a neuronal glutamate transporter. Nature. 1996;383:634–637. - PubMed
-
- Kanner BI, Bendahan A. Binding order of substrates to the sodium and potassium ion coupled l-glutamic acid transporter from rat brain. Biochem. 1982;21:6327–6330. - PubMed
-
- Pines G, Kanner BI. Counterflow of l-glutamate in plasma membrane vesicles and reconstituted preparations from rat brain. Biochem. 1990;29:11209–11214. - PubMed
-
- Kavanaugh MP, Bendahan A, Zerangue N, Zhang Y, Kanner BI. Mutation of an amino acid residue influencing potassium coupling in the glutamate transporter GLT-1 induces obligate exchange. J Biol Chem. 1997;272:1703–1708. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources