The coding of spatial location by single units in the lateral superior olive of the cat. I. Spatial receptive fields in azimuth
- PMID: 11850472
- PMCID: PMC6757576
- DOI: 10.1523/JNEUROSCI.22-04-01454.2002
The coding of spatial location by single units in the lateral superior olive of the cat. I. Spatial receptive fields in azimuth
Abstract
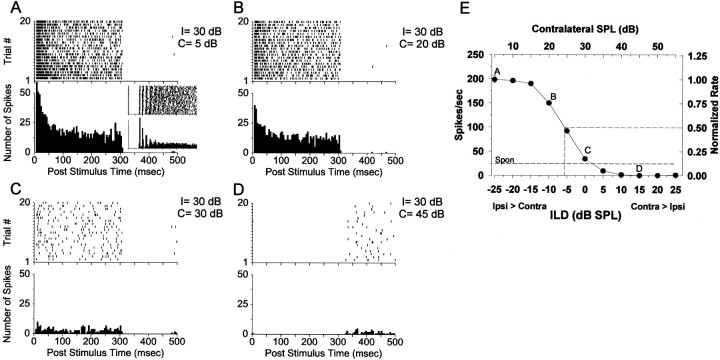
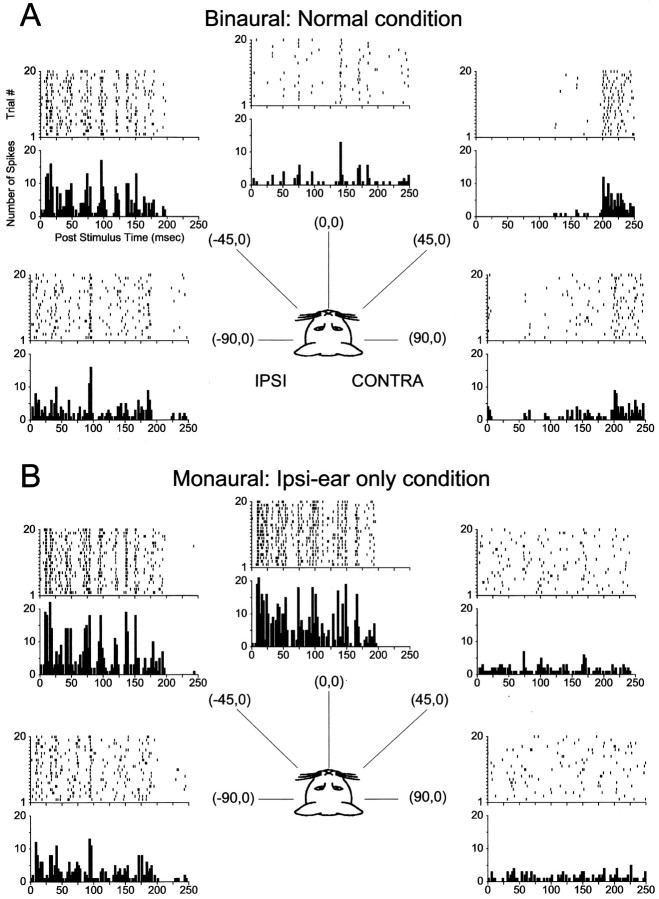
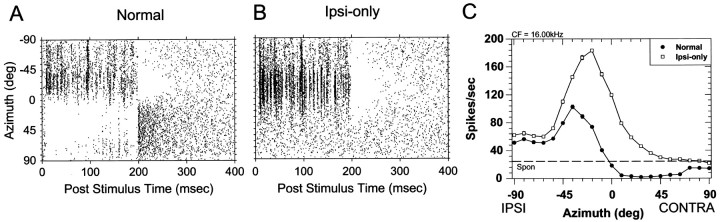
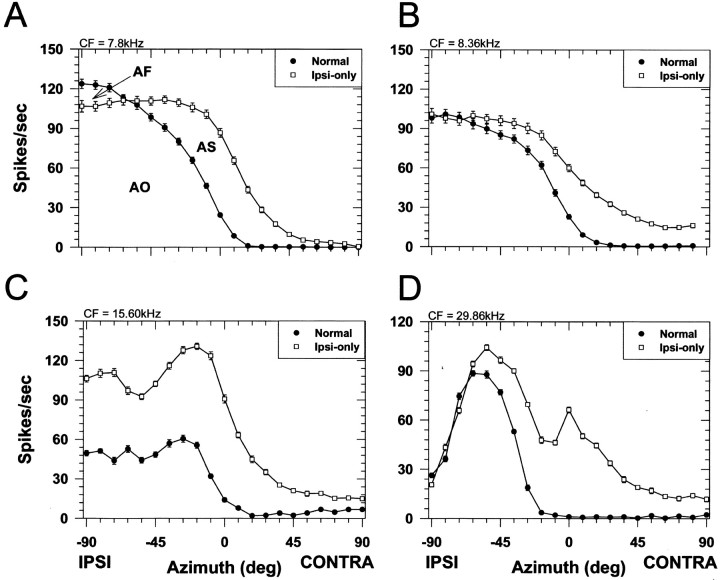
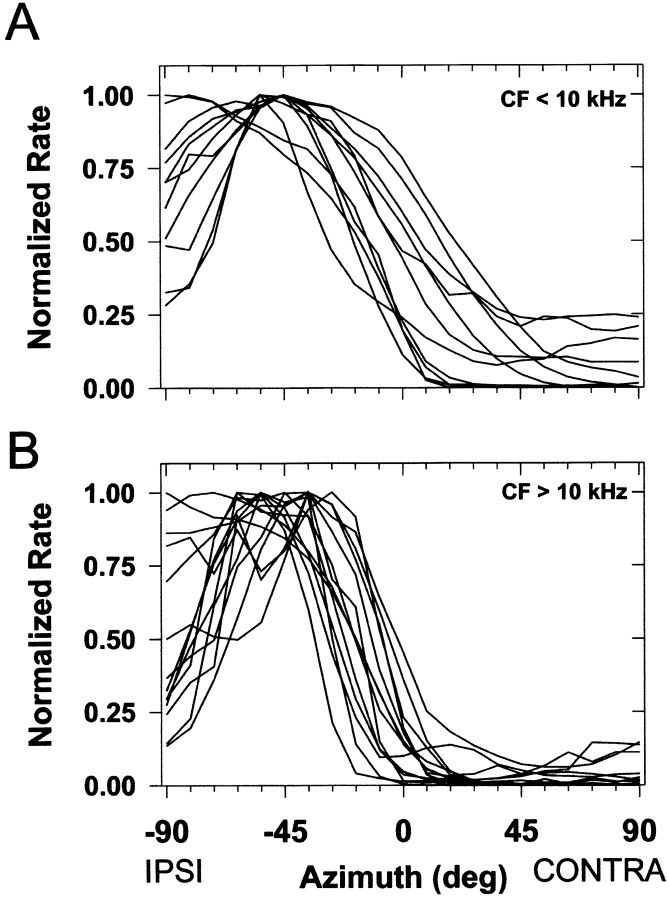
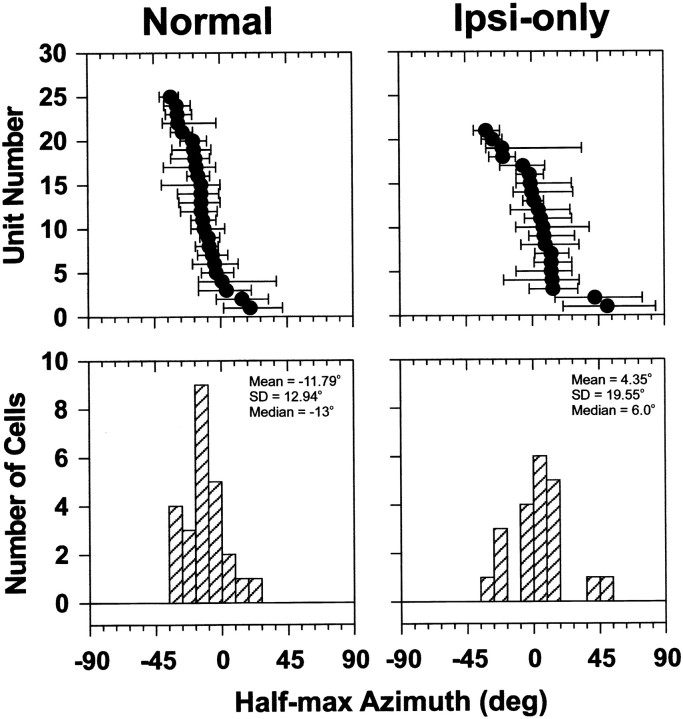
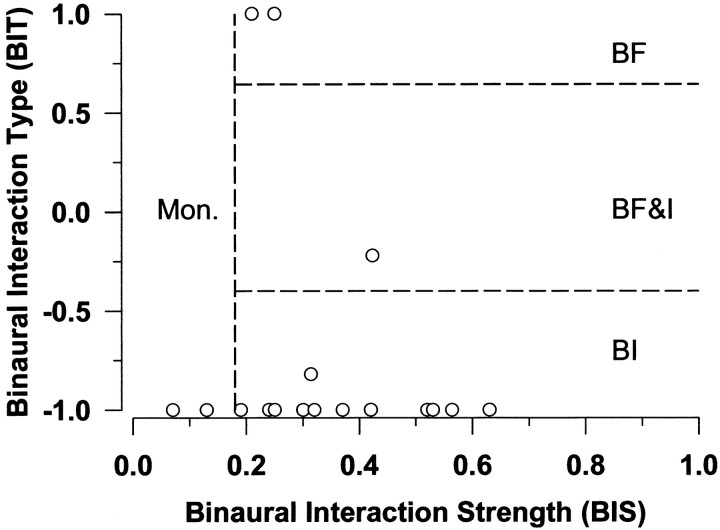
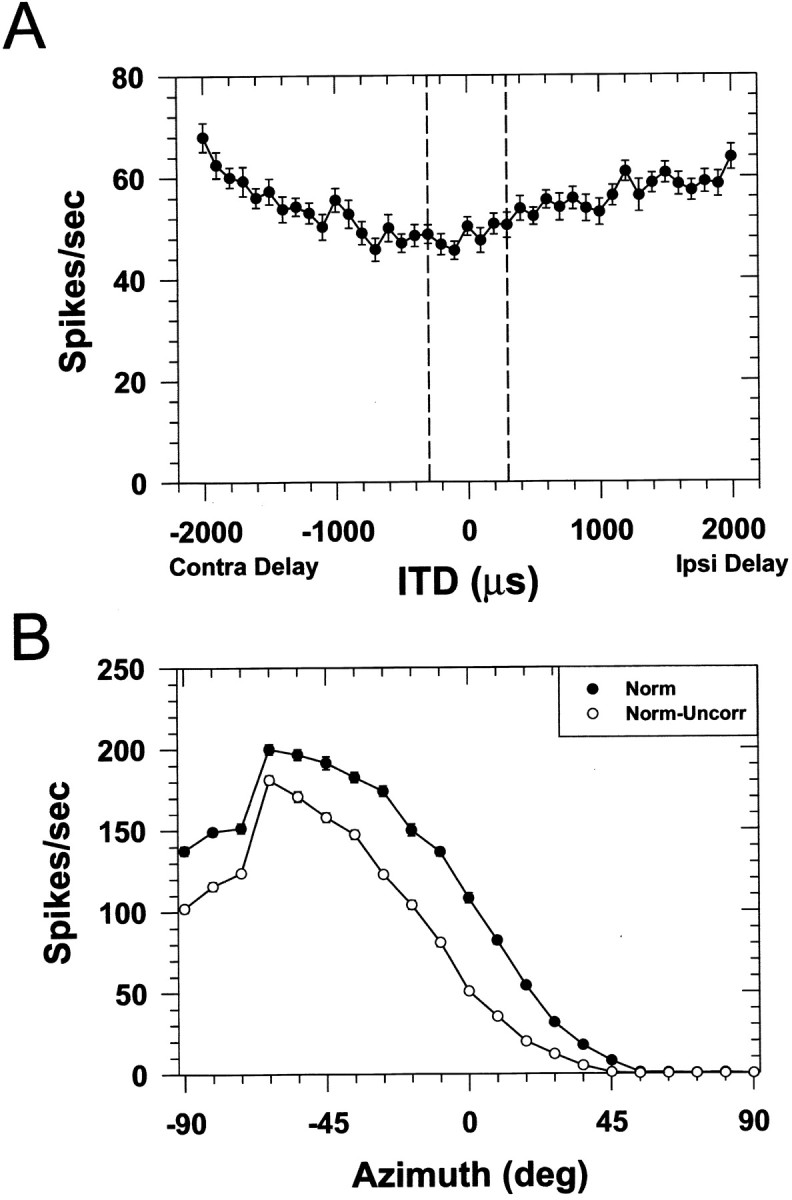
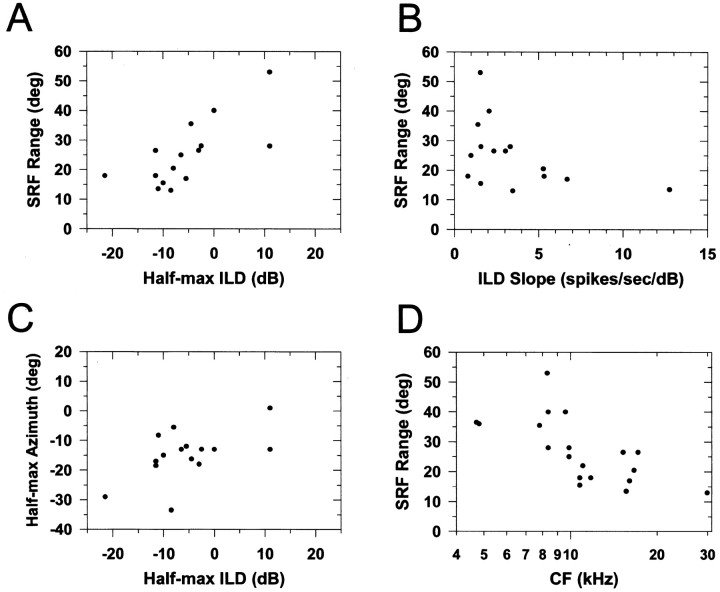
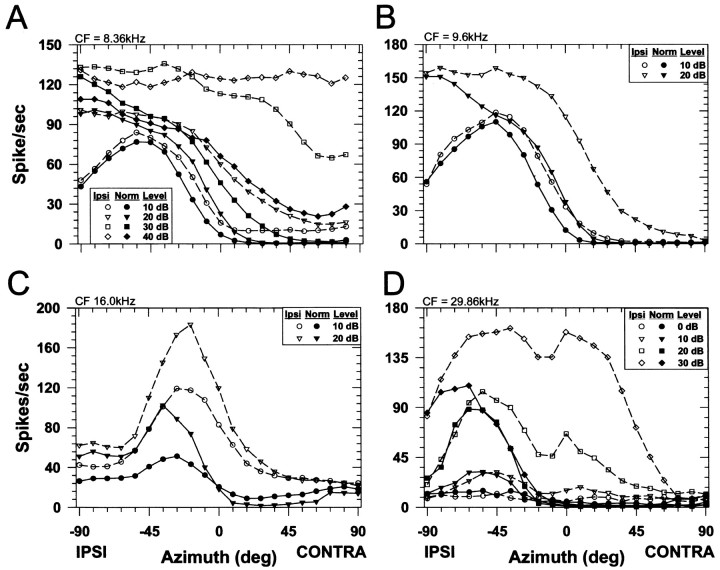
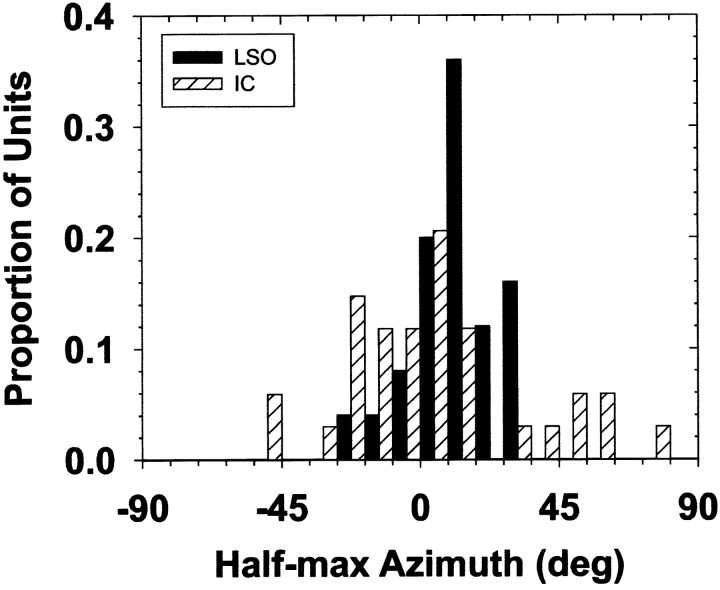
The lateral superior olive (LSO) is one of the most peripheral auditory nuclei receiving inputs from both ears, and LSO neurons are sensitive to interaural level differences (ILDs), one of the primary acoustical cues for sound location. We used the virtual space (VS) technique to present over earphones broadband stimuli containing natural combinations of localization cues as a function of azimuth while recording extracellular responses from single LSO cells. The responses of LSO cells exhibited spatial receptive fields (SRFs) in azimuth consonant with their sensitivity to ILDs of stimuli presented dichotically: high discharge rates for ipsilateral azimuths where stimulus amplitude to the excitatory ear exceeded that to the inhibitory ear, rapidly declining rates near the midline, and low rates for contralateral azimuths where the amplitude to the inhibitory ear exceeded that to the excitatory ear. Relative to binaural stimulation, presentations of the VS stimuli to the ipsilateral ear alone yielded increased rates, particularly in the contralateral field, confirming that the binaural SRFs were shaped by contralateral inhibition. Our finding that LSO neurons respond to azimuth consistent with their ILD sensitivity supports the long-held hypothesis that LSO neurons compute a correlate of the ILD present in free-field stimuli. Only weak correlations between the properties of pure-tone ILD functions and the SRFs were found, indicating that ILD sensitivity measured at only one sound level is not sufficient to predict sensitivity to azimuth. Sensitivity to spatial location was also retained over a wide range of stimulus levels under binaural, but not monaural, conditions.
Figures
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
References
-
- Adam TJ, Schwarz DW, Finlayson PG. Firing properties of chopper and delay neurons in the lateral superior olive of the rat. Exp Brain Res. 1999;124:489–502. - PubMed
-
- Adams JC, Mugnaini E. Dorsal nucleus of the lateral lemniscus: a nucleus of GABAergic projection neurons. Brain Res Bull. 1984;13:585–590. - PubMed
-
- Aitkin L, Jones R. Azimuthal processing in the posterior auditory thalamus of cats. Neurosci Lett. 1992;142:81–84. - PubMed
-
- Aitkin LM, Martin RL. The representation of stimulus azimuth by high best-frequency azimuth-selective neurons in the central nucleus of the inferior colliculus of the cat. J Neurophysiol. 1987;57:1185–1200. - PubMed
-
- Altshuler MW, Comalli PE. Effect of stimulus intensity and frequency on median horizontal plane sound localization. J Aud Res. 1975;15:262–265.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous